


Historia dos bosques de teixo no Norte da Península Ibérica
16-12-2020
A terra ao longo do tempo sufriu numerosas transformacións que condicionaron a vida sobre o planeta. As principais alteracións na historia da terra, foron a deriva continental e o cambio climático. A forza máis importante do cambio climático no pasado era a modificación dos ciclos orbitais da terra, que en último termo determinan a cantidade de insolación que recibe cada parte do planeta e polo tanto o clima nas distintas zonas do globo. Non obstante, os últimos anos da historia da terra, están marcadas polas modificacións derivadas da alteración humana nos ecosistemas naturais.
A información paleoecolóxica permítenos coñecer a evolución do clima e a súa influencia sobre os diferentes biomas e especies. Neste sentido, o estudo dos restos vexetais proporciona unha das visións máis precisas da dinámica dos ecosistemas e as especies. As análises paleobotánicas permiten analizar a resposta das diferentes especies fronte aos cambios climáticos ou outras perturbacións.
A partir do estudo dos macrofósiles (restos de troncos, ramas, follas, froitos, etc.) e microfósiles (grans de pole) das especies arbóreas, pode reconstruírse a súa evolución nun territorio determinado, observando como as diferentes alteracións naturais e antrópicas influíron na súa distribución ao longo do tempo e avaliando os fenómenos de migración, refuxio, extinción, etc.
Aparición das Taxaceae
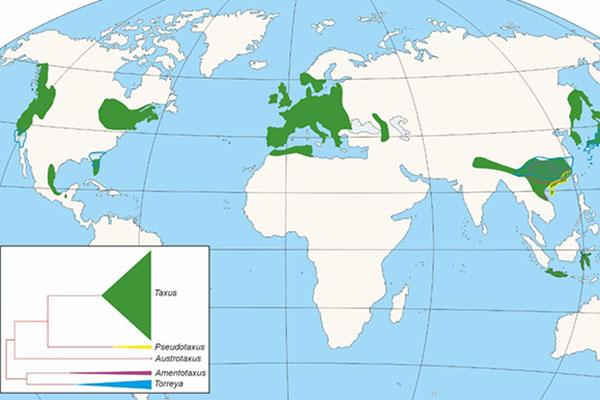
Dentro das ximnospermas, a familia Taxaceae constitúe un grupo claramente diferenciado do resto das coníferas, estando representado na actualidade por cinco xéneros divididos evolutivamente en dous clados, un con tres xéneros Taxus, Austrotaxus e Pseudotaxus, e o segundo con dous (Torreya e Amentotaxus), que en conxunto reúnen unhas 28 especies distribuídas en Eurasia, Norte de África, Nova Caledonia e América (Figura 1), ás que hai que unir distintas especies extintas.
A información paleoecolóxica permítenos coñecer a evolución do clima e a súa influencia sobre os diferentes biomas e especies. Neste sentido, o estudo dos restos vexetais proporciona unha das visións máis precisas da dinámica dos ecosistemas e as especies. As análises paleobotánicas permiten analizar a resposta das diferentes especies fronte aos cambios climáticos ou outras perturbacións.
A partir do estudo dos macrofósiles (restos de troncos, ramas, follas, froitos, etc.) e microfósiles (grans de pole) das especies arbóreas, pode reconstruírse a súa evolución nun territorio determinado, observando como as diferentes alteracións naturais e antrópicas influíron na súa distribución ao longo do tempo e avaliando os fenómenos de migración, refuxio, extinción, etc.
Aparición das Taxaceae
Dentro das ximnospermas, a familia Taxaceae constitúe un grupo claramente diferenciado do resto das coníferas, estando representado na actualidade por cinco xéneros divididos evolutivamente en dous clados, un con tres xéneros Taxus, Austrotaxus e Pseudotaxus, e o segundo con dous (Torreya e Amentotaxus), que en conxunto reúnen unhas 28 especies distribuídas en Eurasia, Norte de África, Nova Caledonia e América (Figura 1), ás que hai que unir distintas especies extintas.
Figura 1. Distribución dos xéneros da Familia Taxaceae e cladograma mostrando a aparición e importancia de cada un deles.
A aparición da familia das Taxaceae sitúase no Mesozoico (Figura 2), concretamente no período Triásico, hai uns 250-200 Ma (Ma: millóns de anos). Taxaceae irradiou dende o sueste de China, diverxendo en varios clados que incluirían a liñaxe europea (Figura 1). O fósil de maior antigüidade de Taxaceae correspóndese cun fragmento dunha rama con follas de Paleotaxus rediviva recuperado en 1908 no sur de Suecia a partires dunha facies de 200 Ma de antigüidade (Nathorst 1908). A partir de Paleotaxus rediviva, evolucionaría durante o Xurásico, hai 140 Ma, o grupo Taxus jurassica (Figura 2). Posteriormente, durante o Cretácico e Cenozoico, diferéncianse novas especies, nun proceso que estivo modulado por importantes cambios tanto na delimitación das unidades continentais, coma no clima, que afectaron á súa distribución e ao illamento de distintos grupos poboacionais en distintos territorios do hemisferio Norte.
A aparición da familia das Taxaceae sitúase no Mesozoico (Figura 2), concretamente no período Triásico, hai uns 250-200 Ma (Ma: millóns de anos). Taxaceae irradiou dende o sueste de China, diverxendo en varios clados que incluirían a liñaxe europea (Figura 1). O fósil de maior antigüidade de Taxaceae correspóndese cun fragmento dunha rama con follas de Paleotaxus rediviva recuperado en 1908 no sur de Suecia a partires dunha facies de 200 Ma de antigüidade (Nathorst 1908). A partir de Paleotaxus rediviva, evolucionaría durante o Xurásico, hai 140 Ma, o grupo Taxus jurassica (Figura 2). Posteriormente, durante o Cretácico e Cenozoico, diferéncianse novas especies, nun proceso que estivo modulado por importantes cambios tanto na delimitación das unidades continentais, coma no clima, que afectaron á súa distribución e ao illamento de distintos grupos poboacionais en distintos territorios do hemisferio Norte.
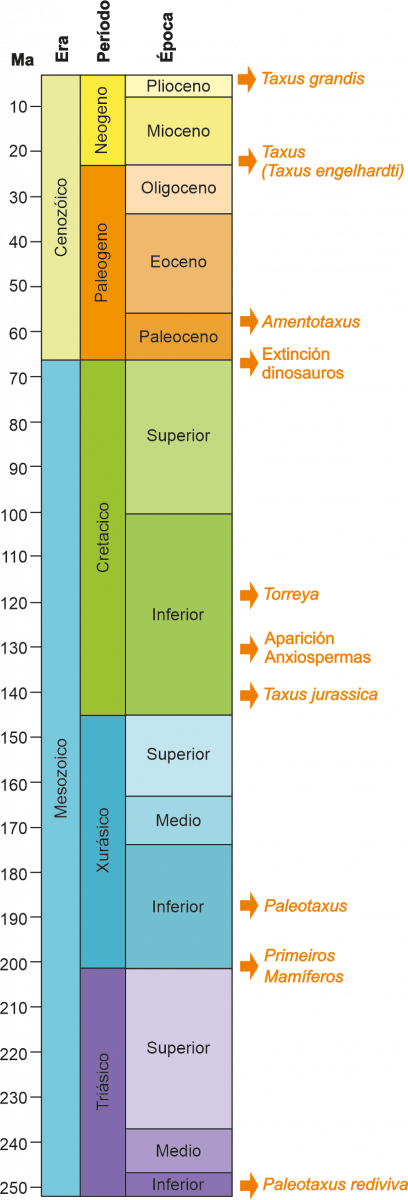
Figura 2. - Localización na escala xeolóxica dos diferentes eventos relacionados coa aparición de Taxus. Ademais, móstranse outros eventos significativos. Ma: millóns de anos.
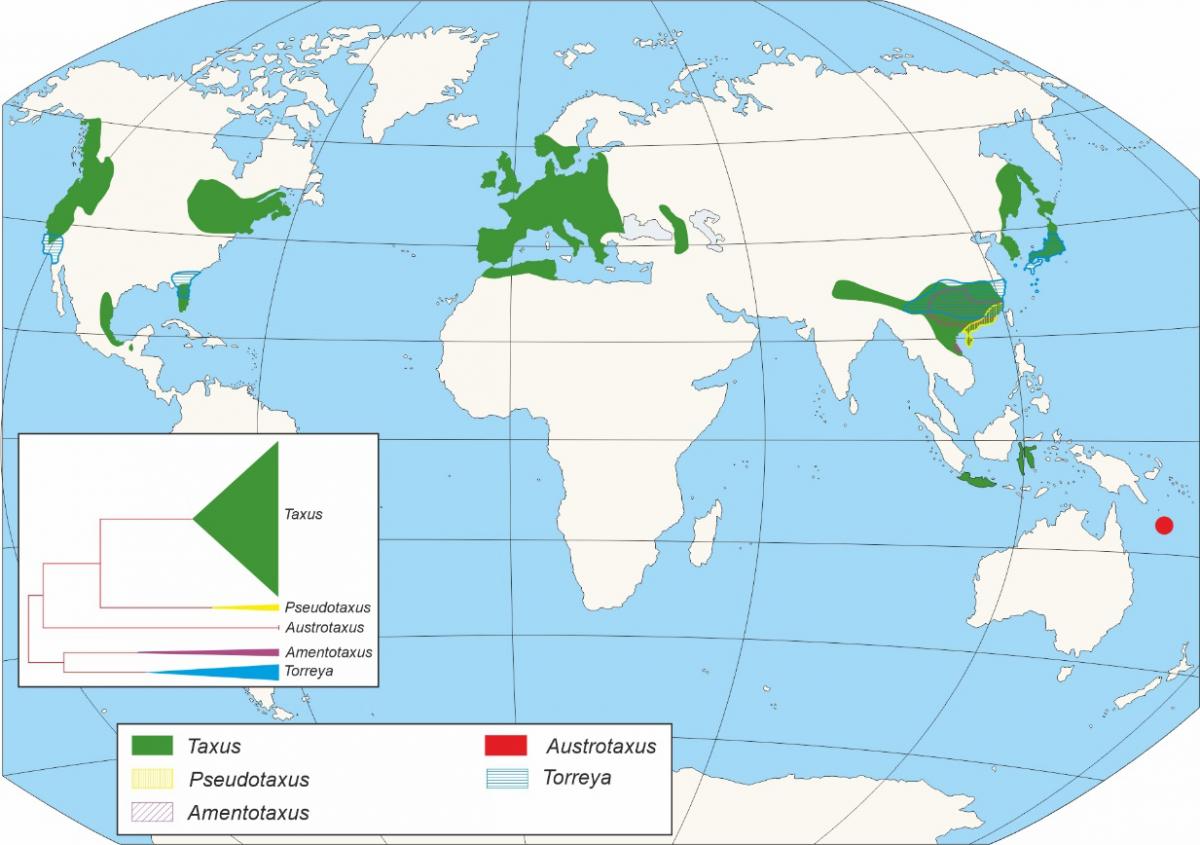
O xénero Taxus (Figuras 2 e 3) está conformado por árbores dioicas con ramificación irregular, con follas aciculares e xemas escamadas sendo o de maior importancia da familia Taxaceae. O xénero Taxus está integrado cando menos por dez especies, aínda que existe un debate en canto ao número das mesmas, principalmente no continente asiático, considerándose unha cuestión aínda non resolta (Hao et al., 2008). Os primeiros restos fósiles adscritos ao xénero Taxus, foron designados como Taxus jurassica (140 Ma). As características morfolóxicas destes permiten a súa asimilación con distintas especies actuais: T. baccata, T. cuspidata, T. brevifolia (Florin, 1958).
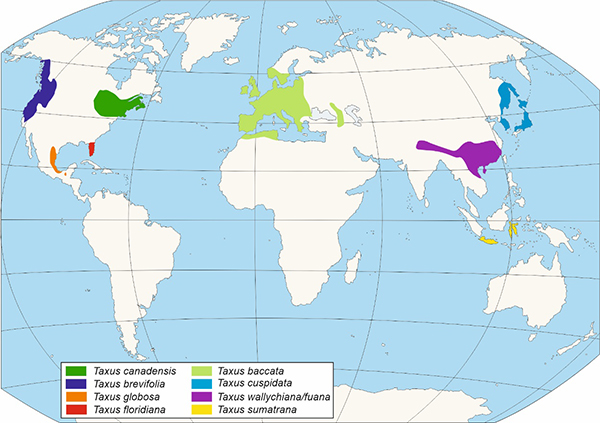
Figura 3. - Distribución das distintas especies do Xénero Taxus segundo Li et al., 2001.
.
.
A era Mesozoica (Figura 2) esténdese entre hai 245 Ma, ata hai 65 Ma. Durante a mesma, a masa continental que constituía o supercontinente Pangea permanece estable sen sufrir grandes alteracións. As condicións climáticas permaneceron basicamente estables, predominando ao longo do continente os ambientes tropicais e subtropicais que marcarían o predominio dos taxa de distribución cosmopolita. As plantas con flores dominantes durante esta era son as ximnospermas, entre as cales se incluirían Taxaceae e outros coníferos (como os ginkgos e as cycas), mentres que, na parte final do Mesozoico, a partir do cretácico inferior, comezan a aparecer os primeiros fósiles de anxiospermas (Figura 2).
Tendo en conta os datos paleobotánicos, Taxus é o xénero de árbores máis antigo do continente europeo, considerándose un xénero relicto. O fósil de Taxus de maior antigüidade, recuperado en Europa, sitúase nos inicios do Mioceno, hai uns 23 Ma (Figura 2). Correspóndese cunhas follas de Taxus engelhardtii localizadas na localidade de Wiesa en Saxonia (Kunzmann & Mai, 2005). O inicio do Mioceno determina o inicio dunha longa etapa de inestabilidade climática a nivel planetario, que marcará a redución dos bosques e especies tropicais dominantes e o paulatino incremento de importancia dos elementos arctoterciarios e esclerófilos. En relación coa transición entre Mioceno e Plioceno, recuperáronse diferentes macrofósiles, incluíndo sementes, ramiñas ou follas, en localidades dispersas do Centro e Leste de Europa.
Na Península Ibérica, os restos fósiles máis antigos correspóndense con grans de pole recuperados en diversas secuencias obtidas dentro da cunca lacustre Plio-pleistocena de Banyoles-Besalú, en Girona (Leroy, 2008) e nas cuncas mineiras de Galicia, Norte e Centro de Portugal (Ramil-Rego et al., 2011). Nestes ámbitos deposicionais, rexístranse algúns grans de pole illados de Taxus ou Taxaceae, relacionados coa época Pliocena (hai 5 millóns de anos)
Tendo en conta os datos paleobotánicos, Taxus é o xénero de árbores máis antigo do continente europeo, considerándose un xénero relicto. O fósil de Taxus de maior antigüidade, recuperado en Europa, sitúase nos inicios do Mioceno, hai uns 23 Ma (Figura 2). Correspóndese cunhas follas de Taxus engelhardtii localizadas na localidade de Wiesa en Saxonia (Kunzmann & Mai, 2005). O inicio do Mioceno determina o inicio dunha longa etapa de inestabilidade climática a nivel planetario, que marcará a redución dos bosques e especies tropicais dominantes e o paulatino incremento de importancia dos elementos arctoterciarios e esclerófilos. En relación coa transición entre Mioceno e Plioceno, recuperáronse diferentes macrofósiles, incluíndo sementes, ramiñas ou follas, en localidades dispersas do Centro e Leste de Europa.
Na Península Ibérica, os restos fósiles máis antigos correspóndense con grans de pole recuperados en diversas secuencias obtidas dentro da cunca lacustre Plio-pleistocena de Banyoles-Besalú, en Girona (Leroy, 2008) e nas cuncas mineiras de Galicia, Norte e Centro de Portugal (Ramil-Rego et al., 2011). Nestes ámbitos deposicionais, rexístranse algúns grans de pole illados de Taxus ou Taxaceae, relacionados coa época Pliocena (hai 5 millóns de anos)
 Figura 4. - Tronco fósil de ximnosperma (aprox. 150 millóns de anos), recollida polo Profesor Montenegro de Andrade en Caldas da Rainha (Portugal) e exposto no edificio do reitorado da Universidade do Porto.
Figura 4. - Tronco fósil de ximnosperma (aprox. 150 millóns de anos), recollida polo Profesor Montenegro de Andrade en Caldas da Rainha (Portugal) e exposto no edificio do reitorado da Universidade do Porto.O clima plioceno, caracterízase pola alternancia de períodos fríos de abundantes chuvias, que nas latitudes máis altas se traducirían en períodos moi fríos con acumulación de xeo, e períodos secos e cálidos. Os rexistros de flora arbórea para este período na Península Ibérica caracterízanse polo dominio absoluto das anxiospermas arctoterciarias, cun grande desenvolvemento das ximnospermas, entre as que se atoparía Taxus, e a presenza dun exiguo grupo de elementos tropicais, entre os que se inclúen distintas especies de palmeiras.
Dinámica de Taxus durante o Plistoceno
O inicio do Plistoceno, hai 2,4 Ma, dá paso a un clima marcado pola sucesión de ciclos de avance glaciar, de aproximadamente 100.000 anos de duración, intercalados con fases de maior benignidade, duns 10.000 anos de duración. Deste modo, durante este dilatado período prodúcese un desenvolvemento cíclico da vexetación en resposta aos cambios climáticos de carácter recorrente. Actualmente, recoñécese que este encadeamento de oscilacións climáticas xogaron un papel determinante na actual distribución das especies arbóreas europeas e dos seus patróns xenéticos.
O estudo da dinámica de Taxus durante a sucesión dos ciclos glaciares baséase fundamentalmente na análise de sondaxes polínicas obtidas en sedimentos de lagos ou turbeiras. Os datos dispoñibles no continente europeo para os interglaciares do Plistoceno inicial redúcense a escasas e dispersas secuencias fragmentadas. Para os últimos ciclos glaciar-interglaciar a información dispoñible amplíase coa presenza de secuencias continuas que aínda son escasas e presentan unha desigual distribución ao longo do continente (Figura 6).
A información paleobotánica dispoñible para o Plistoceno revela que Taxus está presente en Europa dende o inicio deste período. Neste sentido, o taxón fósil cuaternario Taxus grandis, con preto dun millón de anos de antigüidade, é idéntico nas súas características a Taxus baccata (Hartzell, 1991). Ademais, os datos polínicos obtidos en diversos sedimentos atribuídos ás interglaciares iniciais do Plistoceno amosan a presenza de pole de Taxus dende o Plistoceno Inicial, concretamente dende o interglaciar Tiglian-C5, hai 1,8 millóns de anos, (Kase, 1988; Deforce & Bastiaens, 2007).
As secuencias polínicas dispoñibles para o conxunto de Europa continental e as illas británicas mostran un progresivo incremento da importancia deste elemento a medida que transcorren os sucesivos interglaciares do Plistoceno medio, alcanzando a súa máxima importancia paisaxística durante o interglaciar Holsteinian (Jessen et al., 1959, West, 1962, Kelly, 1964, Godwin, 1975, Watts, 1985, Coxon, 2017), hai 400-367 ka (ka: milleiros de anos). En relación con este interglaciar sitúase a lanza de madeira de Taxus recuperada en 1911 (Godwin 1975, Oakley et al., 1977) na localidade de Clacton-on-sea (Essex, Reino Unido), que constitúe un dos máis antigos artefactos fabricados en madeira de Europa.
Na Península Ibérica, os restos polínicos plistocenos de maior antigüidade para Taxus correspóndense co rexistro obtido no rexistro sedimentolóxico do lago de Bòbila-Ordis (Leroy, 2008) situado no NE. O rexistro relaciónase cun período interglaciar dos inicios do Plistoceno, aínda que no estudo non se precisa a cal dos períodos interestadiais do Plistoceno Inicial se refire. Outras evidencias da presenza de Taxus na Península Ibérica correspóndense cos grans de pole identificados en coprólitos de hiena recuperados na zona central peninsular e datados para o Plistoceno Medio (Carrión et al., 2006).
O último ciclo Glaciar - interglaciar no Norte da Península Ibérica
Hai aproximadamente 110.000 anos iníciase o último ciclo glaciar-interglaciar (Figura 5), integrado polo interglaciar Eemiense (hai 115-95 ka) e o glaciar Würm (hai 95-15 ka). A información paleobotánica dispoñible para este período en Europa increméntase, adquirindo unha mellor representación territorial, aínda que aínda dispersa ao longo do continente (Figura 6).
Na Península Ibérica, os restos polínicos plistocenos de maior antigüidade para Taxus correspóndense co rexistro obtido no rexistro sedimentolóxico do lago de Bòbila-Ordis (Leroy, 2008) situado no NE. O rexistro relaciónase cun período interglaciar dos inicios do Plistoceno, aínda que no estudo non se precisa a cal dos períodos interestadiais do Plistoceno Inicial se refire. Outras evidencias da presenza de Taxus na Península Ibérica correspóndense cos grans de pole identificados en coprólitos de hiena recuperados na zona central peninsular e datados para o Plistoceno Medio (Carrión et al., 2006).
O último ciclo Glaciar - interglaciar no Norte da Península Ibérica
Hai aproximadamente 110.000 anos iníciase o último ciclo glaciar-interglaciar (Figura 5), integrado polo interglaciar Eemiense (hai 115-95 ka) e o glaciar Würm (hai 95-15 ka). A información paleobotánica dispoñible para este período en Europa increméntase, adquirindo unha mellor representación territorial, aínda que aínda dispersa ao longo do continente (Figura 6).
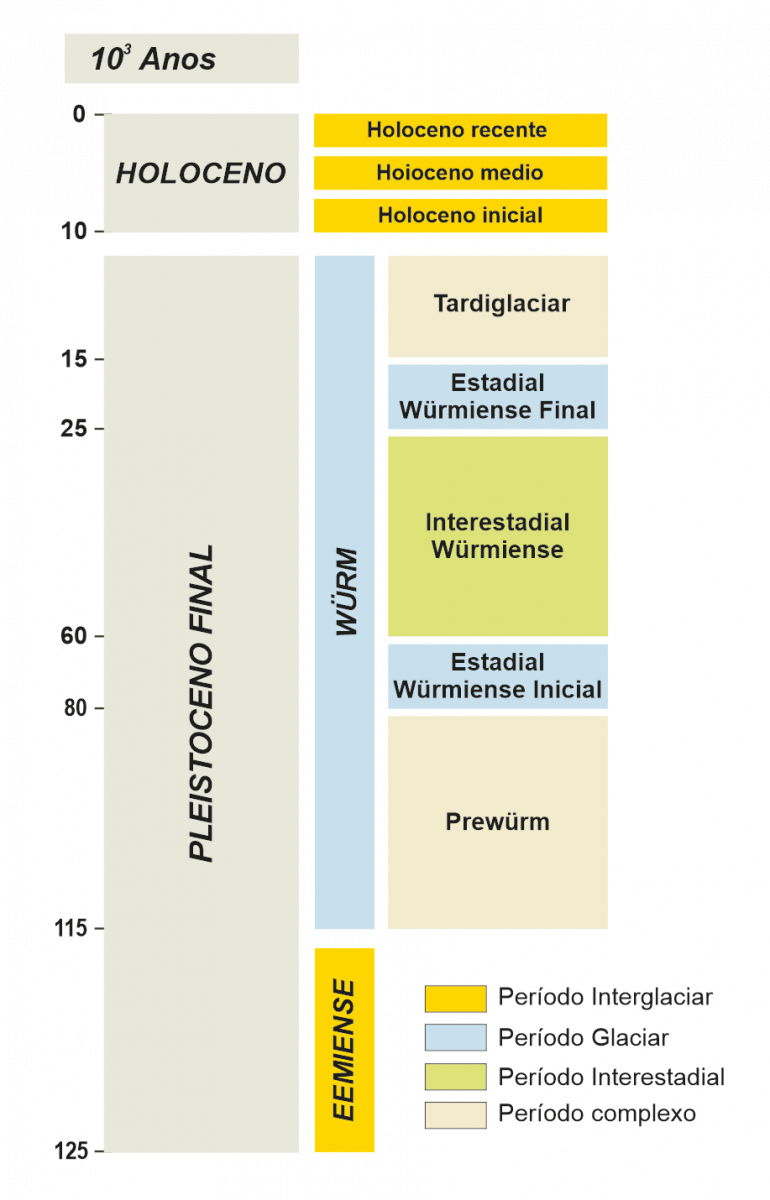
Figura 5. - Periodización do último ciclo glaciar interglaciar no SW de Europa.
Durante o último interglaciar (Eemiense) a grande maioría das secuencias dispoñibles para o continente europeo neste período mostran unha importante fase de expansión de Taxus, principalmente no territorio centro europeo, e diminuíndo a súa importancia cara ao Leste e o Sur, estando mesmo ausente nalgunhas áreas das penínsulas balcánica e itálica. Na Península Ibérica a información paleobotánica para este interglaciar é moi escasa. O único dato que mostra a presenza de Taxus é o rexistro duns grans de pole illados na secuencia Eemiense da turbeira de Padul, en Granada (Camuera et al., 2019).
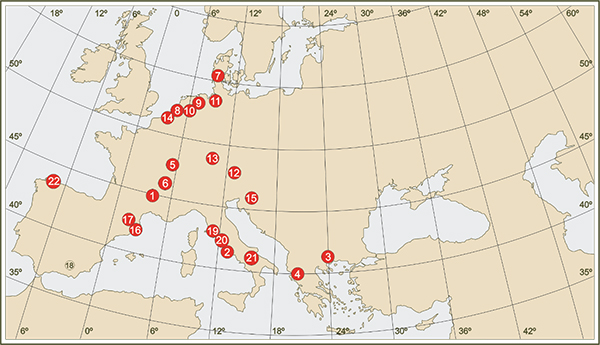
Figura 6. - Localización das principais secuencias polínicas do continente europeo que rexistran máis dun ciclo glaciar do plistoceno (1- Bouchet/Placraux, 2- Valle di Castiglione, 3-Tenaghi Phillipon, 4- Ioannina) ou a totalidade ou grande parte do último ciclo glaciar (5- La Grande Pile, 6- Les Echets, 7- Brorup, 8- Amersfoort, 9- Denekamp, 10- Hengelo, 11- Oerel, 13- Samerberg, 14- Moershoofd, 15- Ljubljana Moor, 16- Banyoles. 17- Pla de l 'Estany, 18- Padul, 19- Lagaccione, 20- Lago di Vico, 21- Lago Grande di Monticchio, 22- Arealonga).
No período comprendido entre hai 95.000-15.000 anos (Figura 5) produciuse no hemisferio Norte un forte arrefriamento do clima que conduciu a importantes cambios na paisaxe e na distribución e evolución das estirpes de flora e fauna silvestre. Os territorios montañosos, xeralmente por enriba dos 900-1.100 m, estiveron afectados por condicións extremadamente frías e dominio dos xeos, incompatibles para a preservación da vexetación arbórea. Por debaixo das áreas cubertas de forma permanente ou temporal polos xeos a paisaxe mantíñase fortemente desarborada. Nas áreas atlánticas establécense formacións herbosas dominadas por gramíneas, mentres que nas continentais a falta de humidade determina o predominio de estepas secas. Por debaixo das formacións herbáceas ou interrelacionadas con estas en distintos mosaicos adaptados ás peculiaridades do territorio se atopan formacións arbustivas de ericoides e leguminosas espiñentas e inermes nas áreas Atlánticas (Queirogais), mentres que nas áreas continentais dominan formacións de Juniperus (Juniperus communis, Juniperus sabina), así como de leguminosas pulvinulares. As formacións herbosas e arbustivas que dominan a paisaxe sustentan unha importante fauna de grandes, medianos e pequenos vertebrados. Os bosques quedan confinados ás áreas máis protexidas do territorio, encontrando de novo unha división entre as áreas Atlánticas con formacións dominadas por anxiospermas caducifolias (Quercus, Alnus, Betula, Corylus, Castanea, Fagus, etc.) e presenza de perennifolias (Ilex, Laurus, Arbutus) e de ximnospermas (Pinus, Abies, Taxus), mentres que nas áreas continentais dominan os Pinus e se reduce a presenza de anxiospermas.
No NW da Península Ibérica, a información paleobotánica para o último período glaciar é moi reducida, limitándose a unha serie de secuencias situadas na actual liña de costa. Os datos dispoñibles non rexistran a presenza de Taxus. A ausencia de rexistro de pole de Taxus durante o glaciar Würm repítese tanto noutras áreas da Península Ibérica como ao longo do continente europeo.
Dinámica de Taxus durante o Tardiglaciar
Nas etapas finais do último ciclo Glaciar-interglaciar prodúcese globalmente un lento aumento da temperatura que repercute na configuración da paisaxe e que desemboca no actual interglaciar, o Holoceno, que se estende entre hai 10.000 anos e a actualidade (Figura 7). Durante este período, denominado Tardiglaciar, prodúcese a fusión dos xeos que cubrían amplas superficies terrestres e mariñas. O dominio dos xeos redúcese, quedando a mediados do Holoceno restrinxido aos cumios de maior altitude. A mellora das condicións climáticas, determinan a expansión do bosque. As formacións arbóreas van ocupando cada vez máis territorio ata alcanzar a súa máxima extensión a mediados do Holoceno, coincidindo co óptimo climático holoceno. No NW da Península Ibérica as formacións caducifolias de anxiospermas fanse dominantes nunha grande parte das áreas máis oceánicas da fachada cantábro-atlántica, mentres que os territorios interiores aparecen dominados por piñeirais ou por mosaicos de piñeirais, bosques de anxiospermas perennifolias e bosques caducifolios.
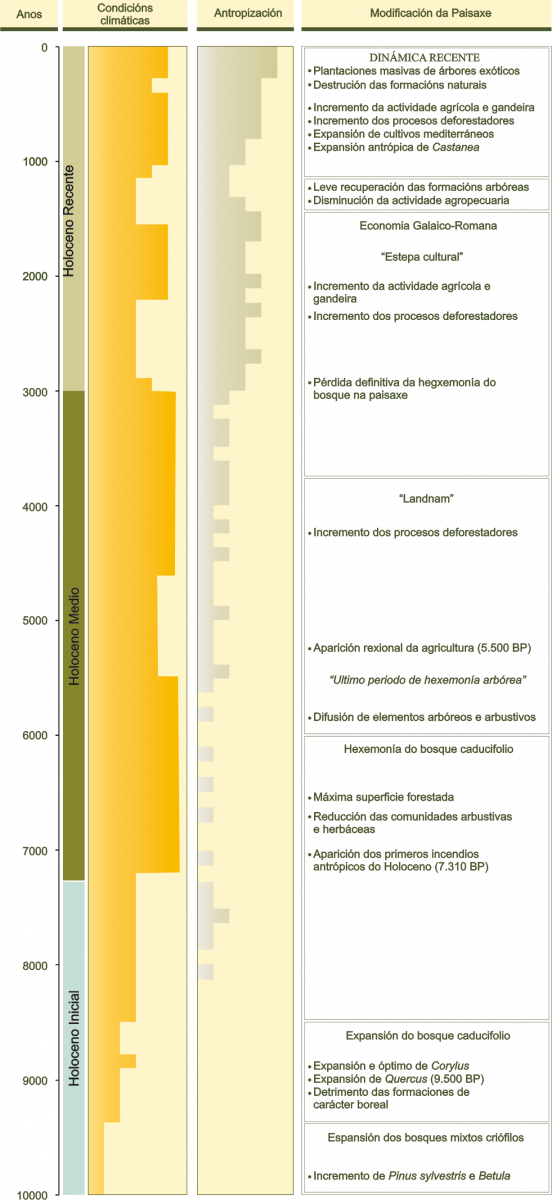
Figura 7. - Periodización ambiental do Holoceno en Galicia.
Durante as fases de deglaciación Tardiglaciar e o inicio do actual interglaciar a escasa presenza de Taxus nos diagramas do NW da Península Ibérica testemuña a persistencia da especie no territorio durante o último ciclo Glaciar-interglaciar, ocupando localidades moi distintas ás actuais, xa que estas últimas na súa maioría mostraban durante o Plistoceno condicións incompatibles co desenvolvemento da especie. Durante este período de desxeo as secuencias de Pena Vella nas serras setentrionais de Galicia (Ramil-Rego, 1992), La Roya (Allen et al., 1986) en Sanabria e Quintanar de la Sierra (Peñalba, 1989) en Burgos, rexistran a presenza de Taxus (Táboa 1, Figura 8).
Dinámica de Taxus durante as fases iniciais do Holoceno
O teixo incrementa no transcurso do Holoceno a súa distribución territorial, progresando en altitude nas distintas rexións do Norte da Península Ibérica (Táboa 1 e Figura 6). Durante as fases de expansión e dominio do bosque que marcan o inicio do Holoceno, aumenta o número de rexistros de Taxus no NW ibérico. Desta forma, ao longo da cornixa cantábrica rexístranse diversas presenzas, tanto de grans de pole en secuencias polínicas, coma de carbóns en depósitos arqueolóxicos (Táboas 1 e 2). En ambos os dous casos, os datos dispoñibles refírense a presenzas puntuais que se corresponderían coa presenza de teixos, como individuos illados incluídos dentro das masas arbóreas dominantes, non detectándose signos da presenza de teixadais.
| Depósito | Tr | HI | HM | HR | Referencia |
| Pena Vella | Ramil Rego, 1992 | ||||
| Pozo do Carballal | Muñoz Sobrino, 2001 | ||||
| La Roya | Allen et al., 1996 | ||||
| Cueto de Avellanosa | Núñez de la Fuente, 2018 | ||||
| Zalama | Pérez Díaz et al., 2016 | ||||
| Los Tornos | Peñalba, 1989 | ||||
| Los Tornos 3-5 | Peñalba, 1989 | ||||
| Quintanar de la Sierra | Peñalba, 1989 | ||||
| Belate | Peñalba, 1989 | ||||
| Atxurri | Peñalba, 1989 | ||||
| Saldropo 1 | Peñalba, 1989 | ||||
| Inurritza 1 | Peñalba, 1989 | ||||
| Fuente del Vaquero | Pérez-Díaz, 2012 |
Táboa 1. - Secuencias polínicas do NW ibérico nas que se identificou a presenza pole de Taxus (Tr: Tardiglaciar, HI: Holoceno Inicial, HM: Holoceno Medio, HR: Holoceno recente).
| Depósito | HI | HM | HR | Referencia |
| Atxoste | Ruíz Alonso, 2003-2007 | |||
| Buruntza | Olaetxea, 1997 | |||
| Ondarre | Mujika et al., 2013 | |||
| Mulisko Gaina | Peñalba, 1987 | |||
| Aizpea | Zapata, 2001 | |||
| Peña Larga | Zapata, 2001 | |||
| Arrubi | Ruiz-Alonso, 2003-2007 | |||
| Mendandia | Iriarte, 2006 | |||
| Peña Parda | Ruiz-Alonso & Zapata, 2003 | |||
| El Mirón | Zapata, 2012 | |||
| Mazaculos | Uzquiano, 1995 | |||
| Aramo II | Beato et al., 2019 | |||
| Peña Oviedo | Carrión Marco, 2005 | |||
| Pala da Vella | Carrión Marco, 2003 |
Táboa 2. - Depósitos do NW ibérico nos que se identificou a presenza de carbóns de Taxus (HI: Holoceno Inicial, HM: Holoceno Medio, HR: Holoceno recente).
Durante as fases iniciais do Holoceno as perturbacións humanas son reducidas e concentradas en determinadas áreas xeográficas, para intensificarse e xeneralizarse posteriormente no Holoceno final, especialmente en territorios como os da Europa occidental, onde se producirá unha forte redución e illamento dos ecosistemas naturais fronte aos antropoecosistemas.
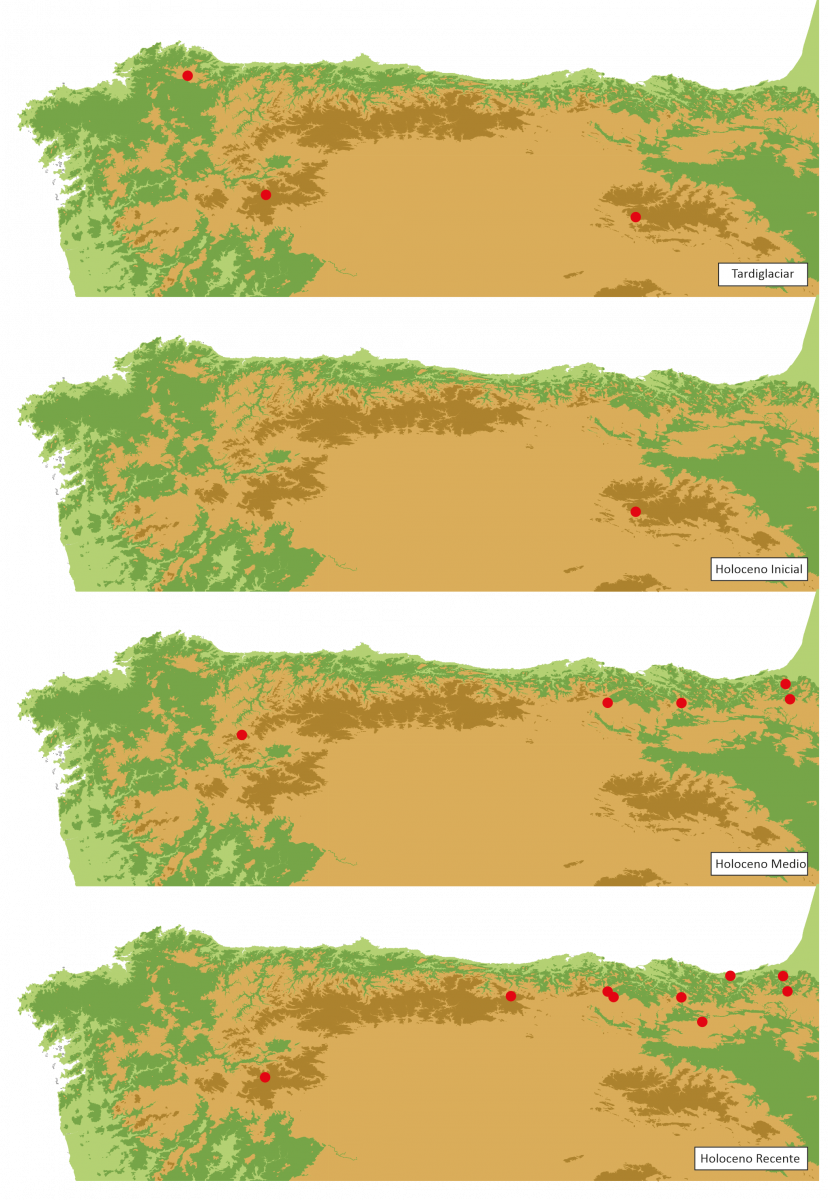
Figura 8. - Secuencias polínicas con presenza de pole de Taxus durante o Tardiglaciar e o Holoceno no NW da Península Ibérica.
Durante o último terzo do Holoceno as secuencias polínicas rexistran a aparición de pole de Taxus nalgunhas secuencias, mentres que noutras se perde o seu rexistro (Figura 8). O rexistro dunha maior cantidade de localidades con pole de Taxus, con respecto a etapas anteriores do Holoceno, relacionaríase coa redución da chuvia polínica das formacións arbóreas dominantes, que enmascararían en moitas secuencias o pole de elementos arbóreos presentes de forma illada entre os bosques maioritarios. Coa redución da importancia paisaxística do bosque, estes elementos minoritarios serían detectados con maior facilidade nas secuencias.
A adopción dos sistemas de produción agrícola e gandeira provocará cambios fundamentais na configuración da paisaxe en toda Europa. No Norte da Península Ibérica este proceso iníciase hai 5.500 anos cando se rexistran os primeiros rexistros de plantas cultivadas. Coincidindo co final da Idade do Bronce e os inicios da Idade do Ferro, as pequenas aldeas de agricultores e gandeiros incrementan a súa produción e a súa pegada sobre a paisaxe. De modo que hai 2.000 anos nas áreas máis intensamente poboadas, os bosques e a vexetación natural viuse desprazada pola configuración de agrosistemas sustentados pola forza das persoas e dos animais domésticos.
O teixo faise agora escaso na paisaxe, e ao igual que outras especies arbóreas queda refuxiado nos enclaves montañosos onde a presión agrícola e gandeira é menos importantes. As paisaxes abertas configuradas por agrosistemas mantéñense e refórzanse durante a Idade Media e a Idade Moderna, incrementando o confinamento das áreas boscosas ás zonas montañosas. En moitos enclaves, especialmente de montaña, o teixo adquire nestas épocas un protagonismo especial vinculado ao seu uso como recurso medicinal, simbólico e espiritual, integrándose nos espazos domésticos e relixiosos, onde permanecen como testemuñas vivas dun complexo e remoto pasado, mentres as formacións nativas se esvaecen e finalmente desaparecen.
As formacións e bosques de teixo, son un compoñente básico da biodiversidade no territorio europeo, forxado ao longo de miles de anos como resultado dos grandes cambios bioxeográficos e climáticos. Bosques e formacións que mantiveron e aínda manteñen en moitas áreas, como acontece na Rexión Atlántica da Península Ibérica, unha intricada relación coas comunidades locais que se beneficiaron dos servizos ecosistémicos que reportan.
Bibliografía
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Beato Bergua, S., Poblete Piedrabuena, M.A., Cunill Artigas, R. (2019). Taxus baccata en la Sierra del Aramo (Macizo Central Asturiano). Boletín de la Asociación de Geógrafos Españoles, 81, 2772, 1–30.
Camuera, J., Jiménez-Moreno, G., Ramos-Román, M.J., García-Alix, A., Toney, J.L., Anderson, R.S., Jiménez-Espejo, F., Bright, J., Webster, C., Yanes, Y., Carrión, J.S. (2019). Vegetation and climate changes during the last two glacial-interglacial cycles in the western Mediterranean: A new long pollen record from Padul (southern Iberian Peninsula),Quaternary Science Reviews, 205: 86-105.
Carrión Marco, Y. (2003). El paisaje vegetal prehistórico de Pala da Vella: primeros resultados antracológicos. In: V. Oliveira Jorge. Contributos das ciencias e das tecnologias para a arqueologia da Península Ibérica: 21-32. UTAD Vila Real, Portugal
Carrión Marco, Y. (2005). El impacto de la economía productora en el paisaje vegetal de Peña Oviedo (Camaleño, Cantabria). In: R. Ontañón Peredo, C. García-Moncó Piñeiro, P. Arias Cabal. Actas del III Congreso del Neolítico en la Península Ibérica: 35-44. Santander.
Carrión, J. S., Scott, L., Arribas, A., Fuentes, N., Gil‐Romera, G. and Montoya, E. (2006). Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. Journal of Quaternary Science, 22: 191–202.
Coxon, P., McCarron, S., Mitchell, F. (Eds) (2017). Advances in Irish Quaternary Studies. Atlantis Advances in Quaternary Science. Atlantis Press, Paris. 316pp.
Deforce, K. & Bastiaens, J. (2007). The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belgian Journal of Botany, 140(2): 222-237.
Florin, R. (1958). On Jurassic taxads and conifers from North-Western Europe and Eastern Greenland. Acta Horti Bergiani Series, 17: 257-402.
Godwin H. (1975). The history of the British Flora. A factual basis for phytogeography. Cambridge University Press, Cambridge. 383 pp.
Hao, D.-C., Huang, B., Yang, L. (2008). Phylogenetic Relationships of the Genus Taxus Inferred from Chloroplast Intergenic Spacer and Nuclear Coding DNA. Biological & pharmaceutical bulletin, 31(2): 260-265.
Hartzell, J.R. (1991). The Yew Tree: A Thousand Whispers. Hulogosi Communications, Oregon, 319 pp.
Heim J. (1970). Les relations entre les spectres polliniques récents et la végétation actuelle en Europe occidentale. Ph.D. thesis, Université catholique de Louvain, Louvain-la-Neuve, Belgium.
Iriarte, M.J. (2006). El entorno vegetal del abrigo de Mendandia y su depósito arqueológico: Análisis palinológico. In: Alday A. (Dir.). El legado arqueologico de Mendandia. Los modos de vida de los últimos cazadores en la Prehistoria de Treviño: 405-418. Junta de Castilla y León. Burgos.
Jessen, K., Andersen, S.Th., Farrington, A. (1959). The interglacial deposit near Gort, Co. Galway, Ireland. Proceedings of the Royal Irish Academy B, 60: 1-77.
Kasse K. (1988). Early-Pleistocene tidal and fluviatile environments in the southern Netherlands and northern Belgium. Ph.D. thesis, Vrije Universiteit, Amsterdam, The Netherlands.
Kelly, M.R. (1964) The Middle Pleistocene of North Birmingham. Philos. T. Roy. Soc. B, 247: 533-592.
Kunzmann L., Mai D.H. (2005) Die Koniferen der Mastixioideen-Flora von Wiesa bei Kamenz (Sachsen, Miozän) unter besonderer Berücksichtigung der Nadelblätter. Palaeontographica B 272(1–6): 67–135.
Leroy, S.A.G. (2008): Vegetation cycles in a disturbed sequence around the Cobb-Mountain subchron in Catalonia (Spain). Journal of Paleolimnology, 40(3), 851-868
Li, J., Davis, C.C., Del Tredici, P., Donoghue, M.J. (2001). Phylogeny and biogeography of Taxus (Taxaceae) inferred from sequences of the internal transcribed spacer region of nuclear ribosomal DNA. Harv. Pap. Bot. ,6:267–274.
Mujika-Alustiza, J.A., Agirre-García, J., Edeso-Fito, J.M., Lopetegi-Galarraga, A., Pérez-Díaz, S., Ruiz-Alonso, M., Tarriño-Vinagre, A., Yusta-Arnal, I. (2013). La continuidad de la actividad pastoril durante la época romana en la zona de Argarbi (sierra de Aralar - Gipuzkoa). Kobie Serie PaleoantroPología, 32: 217-258
Muñoz Sobrino, C. (2001). Cambio climático y dinámica del paisaje en las montañas del noroeste de la Península Ibérica. Tesis Doctoral. Escuela Politécnica Superior. Universidad de Santiago de Compostela.
Nathorst A.G. (1908). Palaeobotanische Mitteillungen.7. Über Palyssya, Stachyotaxus und Paleotaxus. Ksvenka Vetensk Akad Handl 8: 1–16.
Nuñez de la Fuente, S. (2018). Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje atlántico montañoso: la región cantábrica durante el Holoceno. Tesis doctoral. Universidad de Cantabria.
Peñalba Garmendia, M.C. (1989). Dynamique de Végétation Tardiglaciaire et Holocene Du Centre-Nord de l’Espagne d’après l’analyse pollinique. These. Université d’Aix, Marseille III.
Pérez-Díaz, S. (2012). El paisaje vegetal durante la Prehistoria Reciente en la vertiente mediterránea de Euskal Herria— Tesis doctoral. Euskal Herriko Univ., Vitoria-Gastei.
Perez-Díaz, S., López-Sáez, J. A., Pontevedra-Pombal, X., Souto-Souto, M. & Galop, D. (2016). 8000 years of vegetation history in the northern Iberian Peninsula inferred from the palaeoenvironmental study of the Zalama ombrotrophic bog (Basque-Cantabrian Mountains, Spain). Boreas, Vol. 45, pp. 658–672.
Ramil Rego P, Gómez-Orellana L, Muñoz Sobrino C, Tereso JP (2011). Dinâmica natural e transformação antrópica das florestas do noroeste ibérico. In Tereso JP, Honrado JP, Pinto AT, Rego FC (Eds.) Florestas do Norte de Portugal: História, Ecologia e Desafios de Gestão. InBio - Rede de Investigação em Biodiversidade e Biologia Evolutiva. Porto. ISBN: 978-989-97418-1-2. Pp 14-54.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ruiz-Alonso, M. & Zapata, L. (2003). Análisis antracológico del yacimiento arqueológico de Peña Parda. Cuad. Arq. Univ. Navarra, 11: 217-252.
Ruiz-Alonso, M. (2003/2007). Madera carbonizada en los fondos de cabaña de Arrubi y Esnaurreta (Sierra de Aralar, Gipuzkoa): vegetación y recursos forestales en la Edad Media. Kobie (Serie Paleoantr.), 27: 131-150.
Uzquiano, P. (1995). L’evolution de la vegetation a l’Holocene inicial dans le nord de l’Espagne a partir de l’étude anthracologique de trois sites archeologique. Quaternaire 6 (2): 77-83.
Van der Knaap, W.O., van Leeuwen, J.F.N. (1997). Late Glacial and early Holocene vegetation succession, altitudinal vegetation zonation, and climatic change in the Serra da Estrela, Portugal. Review of Palaeobotany and Palynology, 97:3-4: 239-285.
Watts, W.A. (1985) Quaternary vegetation cycles.In: Edwards, K.J. & Warren, W.P. (eds.): The Quaternary history of Ireland: 154-185. Academic Press, London.
West, R.G. (1962) A note on Taxus pollen in the Hoxnian Interglacial. New Phytologist, 61: 189-190.
Zapata, L. (2012). Holocene wood charcoal from El Mirón cave. Vegetation and wood use — In: Straus, L.G. & González Morales, M.R. (Eds.). El Miron cave. Cantabrian Spain. The site and its Holocene archaeological record: 174-196. Univ. New Mexico, Alburquerque.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Allen, J.R.M.; Huntley, B. & Watts, W.A. (1996). The vegetation and climate of northwest Iberia over the last 14.000 yr. Journal of Quaternary Science, 11: 125-147.
Beato Bergua, S., Poblete Piedrabuena, M.A., Cunill Artigas, R. (2019). Taxus baccata en la Sierra del Aramo (Macizo Central Asturiano). Boletín de la Asociación de Geógrafos Españoles, 81, 2772, 1–30.
Camuera, J., Jiménez-Moreno, G., Ramos-Román, M.J., García-Alix, A., Toney, J.L., Anderson, R.S., Jiménez-Espejo, F., Bright, J., Webster, C., Yanes, Y., Carrión, J.S. (2019). Vegetation and climate changes during the last two glacial-interglacial cycles in the western Mediterranean: A new long pollen record from Padul (southern Iberian Peninsula),Quaternary Science Reviews, 205: 86-105.
Carrión Marco, Y. (2003). El paisaje vegetal prehistórico de Pala da Vella: primeros resultados antracológicos. In: V. Oliveira Jorge. Contributos das ciencias e das tecnologias para a arqueologia da Península Ibérica: 21-32. UTAD Vila Real, Portugal
Carrión Marco, Y. (2005). El impacto de la economía productora en el paisaje vegetal de Peña Oviedo (Camaleño, Cantabria). In: R. Ontañón Peredo, C. García-Moncó Piñeiro, P. Arias Cabal. Actas del III Congreso del Neolítico en la Península Ibérica: 35-44. Santander.
Carrión, J. S., Scott, L., Arribas, A., Fuentes, N., Gil‐Romera, G. and Montoya, E. (2006). Pleistocene landscapes in central Iberia inferred from pollen analysis of hyena coprolites. Journal of Quaternary Science, 22: 191–202.
Coxon, P., McCarron, S., Mitchell, F. (Eds) (2017). Advances in Irish Quaternary Studies. Atlantis Advances in Quaternary Science. Atlantis Press, Paris. 316pp.
Deforce, K. & Bastiaens, J. (2007). The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belgian Journal of Botany, 140(2): 222-237.
Florin, R. (1958). On Jurassic taxads and conifers from North-Western Europe and Eastern Greenland. Acta Horti Bergiani Series, 17: 257-402.
Godwin H. (1975). The history of the British Flora. A factual basis for phytogeography. Cambridge University Press, Cambridge. 383 pp.
Hao, D.-C., Huang, B., Yang, L. (2008). Phylogenetic Relationships of the Genus Taxus Inferred from Chloroplast Intergenic Spacer and Nuclear Coding DNA. Biological & pharmaceutical bulletin, 31(2): 260-265.
Hartzell, J.R. (1991). The Yew Tree: A Thousand Whispers. Hulogosi Communications, Oregon, 319 pp.
Heim J. (1970). Les relations entre les spectres polliniques récents et la végétation actuelle en Europe occidentale. Ph.D. thesis, Université catholique de Louvain, Louvain-la-Neuve, Belgium.
Iriarte, M.J. (2006). El entorno vegetal del abrigo de Mendandia y su depósito arqueológico: Análisis palinológico. In: Alday A. (Dir.). El legado arqueologico de Mendandia. Los modos de vida de los últimos cazadores en la Prehistoria de Treviño: 405-418. Junta de Castilla y León. Burgos.
Jessen, K., Andersen, S.Th., Farrington, A. (1959). The interglacial deposit near Gort, Co. Galway, Ireland. Proceedings of the Royal Irish Academy B, 60: 1-77.
Kasse K. (1988). Early-Pleistocene tidal and fluviatile environments in the southern Netherlands and northern Belgium. Ph.D. thesis, Vrije Universiteit, Amsterdam, The Netherlands.
Kelly, M.R. (1964) The Middle Pleistocene of North Birmingham. Philos. T. Roy. Soc. B, 247: 533-592.
Kunzmann L., Mai D.H. (2005) Die Koniferen der Mastixioideen-Flora von Wiesa bei Kamenz (Sachsen, Miozän) unter besonderer Berücksichtigung der Nadelblätter. Palaeontographica B 272(1–6): 67–135.
Leroy, S.A.G. (2008): Vegetation cycles in a disturbed sequence around the Cobb-Mountain subchron in Catalonia (Spain). Journal of Paleolimnology, 40(3), 851-868
Li, J., Davis, C.C., Del Tredici, P., Donoghue, M.J. (2001). Phylogeny and biogeography of Taxus (Taxaceae) inferred from sequences of the internal transcribed spacer region of nuclear ribosomal DNA. Harv. Pap. Bot. ,6:267–274.
Mujika-Alustiza, J.A., Agirre-García, J., Edeso-Fito, J.M., Lopetegi-Galarraga, A., Pérez-Díaz, S., Ruiz-Alonso, M., Tarriño-Vinagre, A., Yusta-Arnal, I. (2013). La continuidad de la actividad pastoril durante la época romana en la zona de Argarbi (sierra de Aralar - Gipuzkoa). Kobie Serie PaleoantroPología, 32: 217-258
Muñoz Sobrino, C. (2001). Cambio climático y dinámica del paisaje en las montañas del noroeste de la Península Ibérica. Tesis Doctoral. Escuela Politécnica Superior. Universidad de Santiago de Compostela.
Nathorst A.G. (1908). Palaeobotanische Mitteillungen.7. Über Palyssya, Stachyotaxus und Paleotaxus. Ksvenka Vetensk Akad Handl 8: 1–16.
Nuñez de la Fuente, S. (2018). Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje atlántico montañoso: la región cantábrica durante el Holoceno. Tesis doctoral. Universidad de Cantabria.
Peñalba Garmendia, M.C. (1989). Dynamique de Végétation Tardiglaciaire et Holocene Du Centre-Nord de l’Espagne d’après l’analyse pollinique. These. Université d’Aix, Marseille III.
Pérez-Díaz, S. (2012). El paisaje vegetal durante la Prehistoria Reciente en la vertiente mediterránea de Euskal Herria— Tesis doctoral. Euskal Herriko Univ., Vitoria-Gastei.
Perez-Díaz, S., López-Sáez, J. A., Pontevedra-Pombal, X., Souto-Souto, M. & Galop, D. (2016). 8000 years of vegetation history in the northern Iberian Peninsula inferred from the palaeoenvironmental study of the Zalama ombrotrophic bog (Basque-Cantabrian Mountains, Spain). Boreas, Vol. 45, pp. 658–672.
Ramil Rego P, Gómez-Orellana L, Muñoz Sobrino C, Tereso JP (2011). Dinâmica natural e transformação antrópica das florestas do noroeste ibérico. In Tereso JP, Honrado JP, Pinto AT, Rego FC (Eds.) Florestas do Norte de Portugal: História, Ecologia e Desafios de Gestão. InBio - Rede de Investigação em Biodiversidade e Biologia Evolutiva. Porto. ISBN: 978-989-97418-1-2. Pp 14-54.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ramil-Rego, P. (1992). La vegetación cuaternaria de las Sierras Septentrionales de Lugo a través del análisis polínico. Tesis Doctoral. Facultad de Biología. Universidad de Santiago de Compostela.
Ruiz-Alonso, M. & Zapata, L. (2003). Análisis antracológico del yacimiento arqueológico de Peña Parda. Cuad. Arq. Univ. Navarra, 11: 217-252.
Ruiz-Alonso, M. (2003/2007). Madera carbonizada en los fondos de cabaña de Arrubi y Esnaurreta (Sierra de Aralar, Gipuzkoa): vegetación y recursos forestales en la Edad Media. Kobie (Serie Paleoantr.), 27: 131-150.
Uzquiano, P. (1995). L’evolution de la vegetation a l’Holocene inicial dans le nord de l’Espagne a partir de l’étude anthracologique de trois sites archeologique. Quaternaire 6 (2): 77-83.
Van der Knaap, W.O., van Leeuwen, J.F.N. (1997). Late Glacial and early Holocene vegetation succession, altitudinal vegetation zonation, and climatic change in the Serra da Estrela, Portugal. Review of Palaeobotany and Palynology, 97:3-4: 239-285.
Watts, W.A. (1985) Quaternary vegetation cycles.In: Edwards, K.J. & Warren, W.P. (eds.): The Quaternary history of Ireland: 154-185. Academic Press, London.
West, R.G. (1962) A note on Taxus pollen in the Hoxnian Interglacial. New Phytologist, 61: 189-190.
Zapata, L. (2012). Holocene wood charcoal from El Mirón cave. Vegetation and wood use — In: Straus, L.G. & González Morales, M.R. (Eds.). El Miron cave. Cantabrian Spain. The site and its Holocene archaeological record: 174-196. Univ. New Mexico, Alburquerque.